第2章 収縮と割れ
収縮異方性
収縮のメカニズム
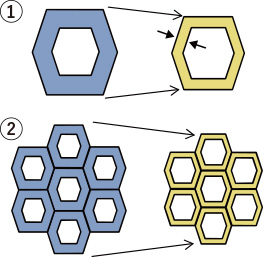 収縮のメカニズムの図
収縮のメカニズムの図木材の乾燥過程では、まず自由水がなくなり、繊維飽和点(含水率約30%)をきると、細胞壁中にある結合水が徐々に減少していきます。結合水が抜けていった細胞壁は、濡れた雑巾が乾くと薄くなるのと同じように、結合水が抜けた量に比例して厚さが減少していきます。細胞壁が薄くなると細胞の径が小さくなり(図中①)、細胞(細胞壁)の集合体である木材が収縮することになります(図中②)。
収縮異方性
 木材の収縮の比率
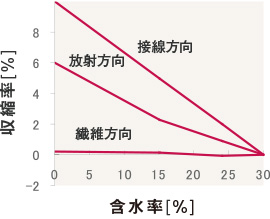
木材の収縮の比率板目方向の収縮率が最も大きく、まさ目方向の約2倍です。長さ方向の収縮率は小さく横方向の10分の1以下です。
繊維飽和点以下になると木材は収縮していきますが、収縮の程度は一様ではなく、方向により異なっています。
収縮率の比は、板目(接線)方向:まさ目(放射)方向:軸方向 = 10:5:0.5~1程度です。つまり、横方向の収縮率が軸方向のそれよりも10倍以上も大きいのです。また、板目方向の収縮率はまさ目の2倍ほどあり、乾燥後の変形や割れが生じる主な原因となっています。
この方向により収縮率が異なる性質は、収縮異方性(いほうせい)と呼ばれます。これは、細胞の並び方や細胞壁内の微細な構造などに起因する性質と考えられています。
| ポイント12 | 細胞壁からの結合水の消失が木材の収縮の始まり。 |
| ポイント13 | 木材の収縮率は方向により異なる(収縮異方性)。 |
| ポイント14 | 収縮の比率は、おおよそ板目:まさ目:長さ=10:5:0.5~1程度。 |
収縮率
収縮率
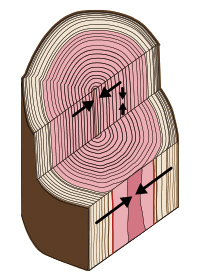
 収縮経過の模式図(木材感想のすべてより)
収縮経過の模式図(木材感想のすべてより)一般に、板目方向は直線か上に凸、まさ目方向は直線か凹、接線方向は僅かに伸びてから収縮する傾向がある。
木材の収縮は含水率の減少にほぼ比例して大きくなります(図参照)。この収縮する割合を収縮率と呼びます。収縮率は様々な樹種について調査されており、全収縮率、気乾収縮率および平均収縮率が発表されています。
気乾収縮率は含水率15%までの収縮率ですので、全収縮率つまり繊維飽和点(約30%)から全乾までの収縮率の半分程度になります。また、気乾収縮率と全収縮率は生材の寸法をもとにした収縮率ですので、挽立寸法にそのまま掛け算して収縮量を求めることができます。
しかし、平均収縮率には少し注意が必要です。平均収縮率は含水率15%から全乾になるまでの含水率 1%当たりの収縮率を含水率15%の寸法をもとに示すことになっています。ですので、全収縮率を1/30にした値に比べ若干大きくなります。
樹種別収縮率
繊維方向の収縮率は比重による差が少なく、全収縮率は約0.2%です。しかし、4m材であれば含水率15%までに4mm程度短くなることに注意が必要です。また横方向の収縮率は、ほぼ比重に比例して大きくなり3~10%の範囲にあります。表に代表的な樹種の収縮率を示しますが、小さな試験片のデータで、実際の材とは多少異なることに注意してください。
| 樹種 | 全乾 比重 | 平均収縮率 | 気乾収縮率 | 全乾収縮率 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 接線 | 放射 | 繊維 | 接線 | 放射 | 繊維 | 接線 | 放射 | 繊維 | ||
| スギ(熊本) | 0.36 | 0.239 | 0.092 | 0.009 | 3.03 | 1.11 | 0.02 | 6.50 | 2.48 | 0.15 |
| スギ(秋田) | 0.33 | 0.259 | 0.093 | 0.011 | 3.46 | 1.07 | 0.03 | 7.19 | 2.44 | 0.19 |
| ヒノキ | 0.37 | 0.205 | 0.106 | 0.013 | 3.45 | 1.49 | 0.05 | 6.43 | 3.07 | 0.25 |
| アカマツ | 0.52 | 0.314 | 0.154 | 0.013 | 4.40 | 1.86 | 0.03 | 8.90 | 4.13 | 0.20 |
| カラマツ | 0.50 | 0.310 | 0.143 | 0.011 | 4.13 | 1.73 | 0.01 | 8.61 | 3.85 | 0.18 |
| トドマツ | 0.39 | 0.375 | 0.120 | 0.010 | 4.14 | 0.96 | 0.03 | 9.53 | 2.75 | 0.19 |
| ポイント15 | 全収縮率、気乾収縮率、平均収縮率が一般に示されている。 |
| ポイント16 | 繊維方向の全収縮率は約0.2%、横方向収縮率は比重に比例する。 |
変形と狂い
木取りによる変形
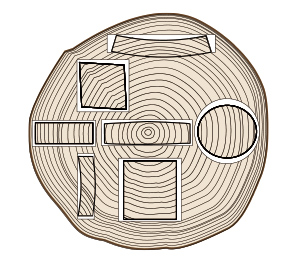 木取りによる断面変形の模式図
木取りによる断面変形の模式図収縮異方性の項目で説明したように、板目方向の収縮率はまさ目方向の2倍ほどあります。また、追いまさ方向ではそれらの中間的な収縮率を示します。ですので、同じサイズの材を製材したとしても、乾燥後には木取りされた丸太の位置によって図のような異なった変形をしてしまうのです。今後、乾燥材生産が前提となった製材の木取りは、乾燥後の変形や収縮を見込んだ寸法に挽くことがポイントになってきます。
狂い
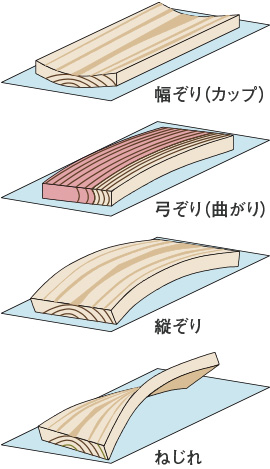
そり、曲がり、ねじれを狂いと言います。
狂いには、正常な材にも生じるものと、欠点によって生じるものがあります。
板目材が木表側に反る幅ぞり(カップ)は、正常な材にも生じるもので、放射方向と接線方向の収縮率が異なることが原因となっています。
まさ目板が辺材側を凹にして曲がることを弓ぞりあるいは単に曲がりと言い、板が厚み方向に反ったものを縦ぞりと呼びます。これらは、幹が太る時に幹の中に貯め込まれた力(成長応力)やあて材の存在に原因される場合が多いと考えられています。
ねじれを含め狂いのほとんどは交錯木理や旋回木理などの細胞配列の乱れによる場合が多く、また節の周囲も狂いがでやすい部分です。
狂いの低減には、低温での圧締乾燥や初期および中間の蒸煮が有効と言われています。
| ポイント17 | 放射方向と接線方向との収縮率の違いにより変形が生じる。 |
| ポイント18 | 成長応力、あて、細胞配列の乱れなどにより狂いは生じる。 |
割れ
初期割れ
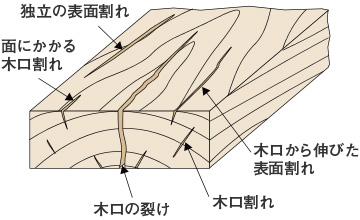 乾燥初期に生じやすい割れ
乾燥初期に生じやすい割れ乾燥初期に生じる割れには、木口割れと表面割れがあります。どちらも木口部分や材表面が先に乾燥して収縮するために起こります。板材などで木口部分を貫通した割れを木口の裂けと呼び、最も問題となる損傷です。
初期割れが発生する原因は、
①水分傾斜、つまり表面は乾いて収縮しようとしても、内部には水分が残ったままで寸法に変化が起こらず、逆に表面には引っ張りの力が働くこと、
②収縮率の差、つまり板目方向の収縮率がまさ目方向の2倍近くあること、
③板目方向の強度が最も弱い(強さは繊維方向>>まさ目方向>板目方向)ことです。
逆に言いますと、収縮率の差が小さい樹種や、横への引張力に耐えられる材であるならば、割れが生じにくいことになります。また、木口割れの多くは、丸太の木口割れが製材後にも残ったために生じると考えられています。
内部割れ
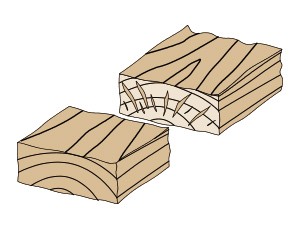 乾燥末期に生じる内部割れ
乾燥末期に生じる内部割れ乾燥後に材を横切りした時に見られる内部の割れを内部割れと言います。この内部割れは感想の末期に発生します。
この割れが発生する原因は、
①初期割れとは逆に、乾燥末期に内部が乾燥し収縮しようとする時に、引張された表層に引っ張られる力に耐えられなくなることや、
②落ち込みと呼ばれる細胞のつぶれが内部に生じた結果だと考えられています。
| ポイント19 | 乾燥初期に発生しやすい割れには、木口割れと表面割れがある。 |
| ポイント20 | 乾燥末期には内部割れが生じる場合がある。 |
特異な収縮
乾燥応力
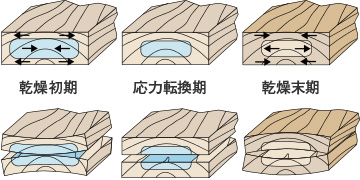 乾燥過程の応力と応力による変形
乾燥過程の応力と応力による変形乾燥初期には、表層は繊維飽和点をき って収縮しようとし、その内部はまだ収縮しません。この時、表層には内部が突っ張るので引張応力が、逆に内部には圧縮応力が働くのです。乾燥がさらに進むと表層は引っ張られたまま固定化し、内部も繊維飽和点をきって収縮しようとしだします。この時、表層の引張力が消え逆に圧縮力が、内部には引張力が働くようになります。この応力が逆転する時期を応力転換期と呼びます。
ドライングセット
薄くて小さな材が乾燥する時には、ほぼ自由に収縮でき、乾燥条件に関係なく収縮率は変わりません。しかし、ある程度の厚材では、単純な収縮以外に、乾燥応力、細胞のつぶれ(落ち込み)、重しの圧力などのいろいろな要素が複雑に絡みあい、乾燥条件や個々の材の質により収縮率に違いが現れます。自由収縮に対して、ある力の働く乾燥条件下での収縮を、ドライングセットと呼びます。
例えば、引張りながら乾燥させた材の収縮率は減少します(引張セット)。事実、同じサイズの材を温度を変えて乾燥してみると、低温乾燥された材の収縮率が若干小さくなる現象が見られます。これは表層に引張応力が、長く働くためだと考えられます。
落ち込み
樹種によって木材が乾燥する過程で、特定の細胞や組織は含水率が繊維飽和点以上でつぶれる場合があります。これを細胞の落ち込みと呼びます。この状態が特定の箇所に集中して発生すると、板にくぼみが生じます。この状態を落ち込みが生じたと言います。また、板全体に発生すると収縮率が大きくなるので、異常収縮とも呼ばれます。細胞の落ち込みの原因の一つは、水の引張力と考えられており、温度が高いと発生しやすくなります。
| ポイント21 | 一般に乾燥過程の材には乾燥応力が働いている。 |
| ポイント22 | 乾燥過程で自由収縮は稀で、多少はドライングセットされる。 |
| ポイント23 | 繊維飽和点以上で細胞がつぶれ、落ち込みが生じることがある。 |




